The Third Week Of Life:
GASTRULATION AND THE FORMATION OF THE TRILAMINAR EMBRYO
Professor Alfred Cuschieri
Department of Anatomy
University of Malta
In most animals the blastula develops into a gastrula that consists of
a distended sac-like structure with an opening at one end. Important changes occur at the opening of
the gastrula involving migration of cells and leading to the formation of a
trilaminar embryo. In humans the embryo
does not form a gastrula, but the changes leading to the formation of the
trilaminar embryo occur and are known as gastrulation.
By the 13th day the embryo has the form of a bilaminar disc
with a distinct polarity.
The amniotic cavity and the yolk sac can be visualised as two
hemispheres, with their apposed surfaces forming the bilaminar embryo.
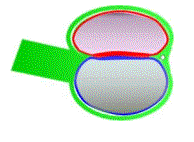
The main changes associated with gastrulation affect the epiblast.
When viewed from above, i.e through the amniotic cavity, the epiblast
appears as an oval disc. The connecting
stalk marks the caudal end of the embryo.
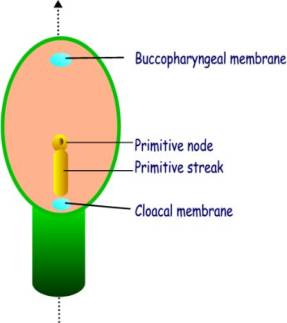
The primitive streak appears in the caudal half of the epiblast, and
lies along the cranio-caudal axis.
It consists of :
·
The primitive node and pit
·
The primitive streak and groove
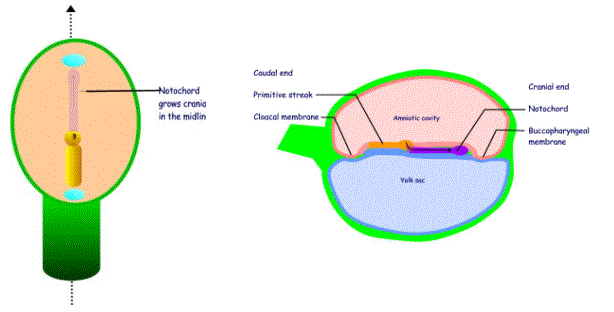
The buccopharyngeal membrane and the cloacal membrane are two areas that mark the future mouth and anus respectively. They are situated in the midline at the cranial and caudal ends respectively.
Three important processes occur at the primitive streak:
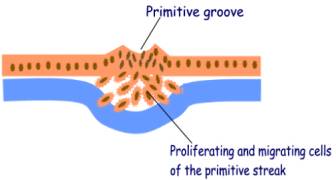
1.
Cell proliferation - causes heaping up of the cells and is the source
of a new layer of cells
2.
Cell migration by amoeboid movement – the cells insinuate
themselves between the epiblast and hypoblast
3. Cell determination - the cells arising from the primitive streak are determined to give rise to different rudiments
The notochord
The notochordal process grows out from the primitive node grows
as a rod of cells that migrate cranially in the midline. Its growth is limited
by the buccopharyngeal membrane. The
most cranial part of the notochord is termed the prochordal plate.
The notochordal process becomes canalised forming a hollow tube
communicating with the primitive pit.
The floor of the tube and the underlying endoderm break down, and a
temporary communication is established between the amniotic cavity and the yolk
sac. This communication is termed the neurenteric
canal. It is very short-lasting as
the notochord proliferates to form a solid cord.
Intra-embryonic mesoderm
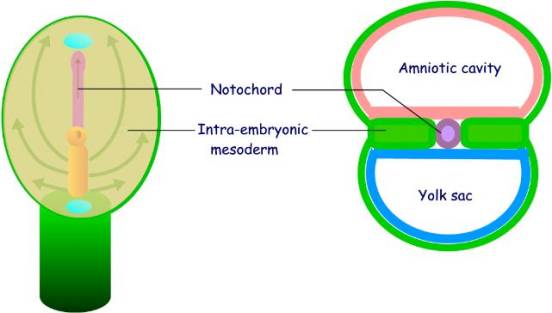 While the cells
from the primitive node proliferate and migrate to form a midline
notochord, cells from the primitve
streak migrate laterally and cranially between the epiblast and
hypoblast to form the intra-embryonic mesoderm. The meosderm lies on either side of the notochord,
the buccopharyngeal membrane, and the
cloacal membrane. The epiblast and
hypoblast remain in contact at the buccopharyngeal and cloacal membranes, which
are not separated by mesoderm.
While the cells
from the primitive node proliferate and migrate to form a midline
notochord, cells from the primitve
streak migrate laterally and cranially between the epiblast and
hypoblast to form the intra-embryonic mesoderm. The meosderm lies on either side of the notochord,
the buccopharyngeal membrane, and the
cloacal membrane. The epiblast and
hypoblast remain in contact at the buccopharyngeal and cloacal membranes, which
are not separated by mesoderm.
Growth of the bilaminar disc.
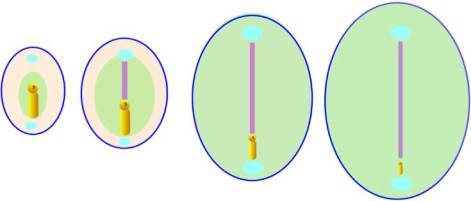
As the intra-embryonic mesoderm spreads out from the primitive streak,
the whole embryo increases in size and the primitive streak becomes relatively
smaller. When the process of
gastrulation is complete the primitive streak disappears.
Changes consequent on gastrulation
With the process of gastrulation the following changes have occurred:
1.
The embryo becomes a trilaminar embryo that is still in the form
of a flat disc.
2.
Epiblast and hypoblast are now known as ectoderm and endoderm
respectively.
3.
Mesoderm does not extend between epiblast and hypoblast at the
buccopharyngeal and cloacal membranes
4.
Intra-embryonic mesoderm merges with the extra-embryonic mesoderm at
the periphery of the embryonic disc
5.
Gastrulation converts the embryo into a trilaminar disc
6.
Gastrulaton can be inhibited by drugs that interfere with actin
filament formation – they inhibit cytokinesis and cell migration
7.
The migrating cells can be traced experimentally in chick embryos by
transplanting embryonic cells of the primitive streak from quail embryos. Quail cells have nuclei that are
morphologically different fromthose of chick cells and can be distinguished
microscopically.
Mesoderm forms several distinct masses:
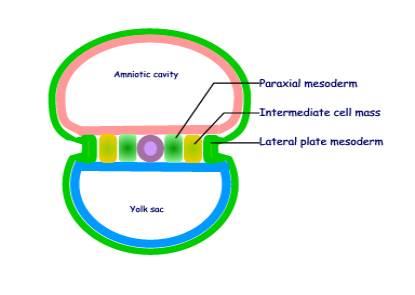
I
Mesoderm in the lateral part of the embryo is
divided into three distinct longitudinal masses:
(a) Paraxial mesoderm
- a longitudinal column of cells that lies next to the notochord
- it gives rise to the axial
skeleton and skeletal muscle
- it becomes segmented
(b) Intermediate cell
mass
- it gives rise to the genitourinary
system
(c) Lateral plate
mesoderm
- gives rise to body wall
structures
- is continuous with the extra-embryonic mesoderm
- splits into two layers
enclosing the intra-embryonic coelom
II
The Septum transversum
- Forms a transverse bar of mesoderm cranial to the buccopharyngeal
membrane
– Is the only place
where mesoderm extends across the midline
– Is the region
where the future heart, diaphphragm and liver will develop.
III
The Caudal mesoderm
- Lies caudaly lateral to the primitive streak
– Is the region
that gives rise to the caudal
structures including the pelvic viscera.
The paraxial mesoderm is segmented
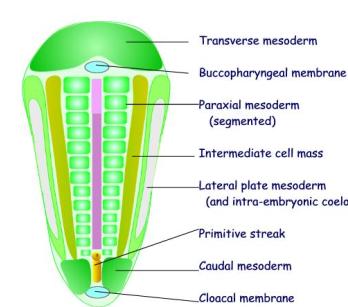 Of the three
blocks pf mesoderm only the paraxial mesoderm is segmented. The segments are termed somites. The first
somite appears on day 20 at the cranial end close to the prochordal plate.
Of the three
blocks pf mesoderm only the paraxial mesoderm is segmented. The segments are termed somites. The first
somite appears on day 20 at the cranial end close to the prochordal plate.
Somites develop in cranio-caudal sequence to form 42 to 44 somites by
day 30.
The somites are:
4 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral,
and 3 coccygeal.
A few other somites at the caudal end degenerate.
A coelom develops in the lateral plate mesoderm.
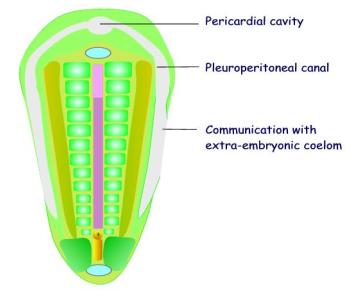 The
intra-embryonic coelom develops in the lateral plate mesoderm and extends into
the transverse mesoderm. It develops by
the breaking down of cells by apoptosis (programmed cell death). The intra-embryonic coelom is shaped like an
inverted U-tube, and is divided into three parts:
The
intra-embryonic coelom develops in the lateral plate mesoderm and extends into
the transverse mesoderm. It develops by
the breaking down of cells by apoptosis (programmed cell death). The intra-embryonic coelom is shaped like an
inverted U-tube, and is divided into three parts:
(a)
The bend is situated in the transverse mesoderm and is dilated to form
the pericardial cavity.
(b)
The stems of the U-tube extend
into the lateral plate mesoderm were it forms the paired pleuro-peritoneal
canals.
(c) The distal (caudal) end of the intra-embryonic coelom communicates laterally with the extra-embryonic coelom.
The Concept of Induction
The embryological processes of development follow a definite
co-ordinated sequence. Induction is 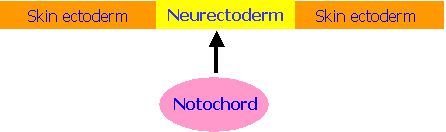 the stimulation
of a group of cells to undergo differentiation by another closely-situated
group of cell
the stimulation
of a group of cells to undergo differentiation by another closely-situated
group of cell
An example of induction is the initial development of the central
nervous system. Here the notochord (a
midline rod of cells) stimulates the overlying ectoderm to differentiate into neurectoderm.
The Neural Plate

The neurectoderm is now termed the neural plate. Like the notochord it is limited craniallly
by the buccopharyngeal membrane and caudally by the cloacal membrane.
The cranial end of the neural plate
·
is broader
than the more caudal part
·
overlies the prochordal plate
·
gives rise to the brain
The main part :
·
Overlies the notochord
·
Gives rise to the spinal cord
The caudal end of the embryo
continues to elongate by growth from the primitive streak
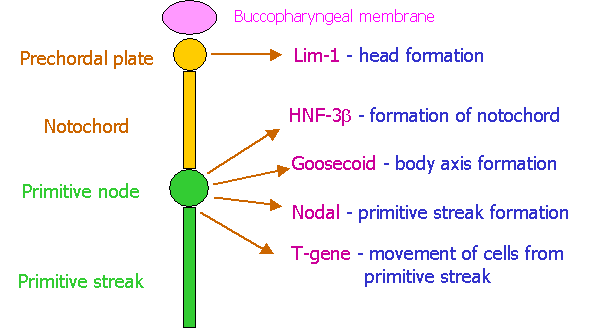
Genetic organization of the Primitive Streak.
The diagram below shows that
primitive node and streak release a number of genetically controlled
factors. These are shown on the right
of the diagram together with their main function in patterning of the embryo. It
is not important to memorise the strange names given to these factors. Note how these factors play a critical role
in determining the cranio-caudal axis of the embryo. The primitive node is the first and central structure. It
determines the growth of the notochord cranially (by factor NHF-3b) and of the primitive streak caudally (by the factor
nodal) and the body axis (by the factor goosecoid). It is also responsible for inducing the
cells to become motile and migrate to pre-determined sites (by the factor T-gene). Note also how the prochordal plate
determines the cranial end of the embryo (by factor Lim-1).
Establishment
of right-left asymmetry.
This is the second critical event in patterning of the embryo, and in
determining the asymmetric development of the viscera on the right and left
sides of the body. This right left
differentiation is determined by several laterality genes, the main ones of
which are show below.
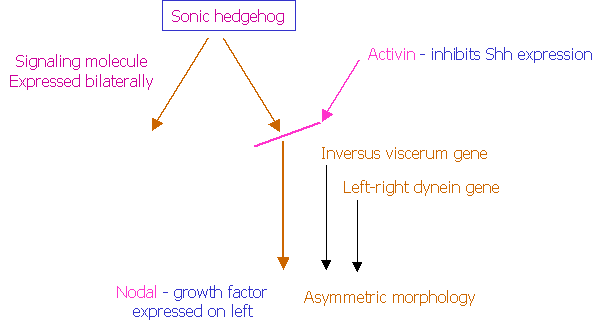
The signalling molecule known as sonic hedgehog (Shh) is expressed
bilaterally and symmetrically. Activin
is expressed only on the left, inhibits the further action of Shh, and causes a
differential expression of the growth factor nodal. There are also genes that
specifically determine development of structures only on the left side. These have been called Inversus viscerum
and left-right dynein. Mutations
of these genes cause “situs inversus” where the apex of the heart, the spleen
and the stomach are situated on the right instead of of the left.
There are three big classes of factors that are involved in the genetic
control of early embryonic development:
1. Transcription factors. These
are
factors that :
·
Act within the cells that produce them
·
Bind to DNA and controls transcription of other genes
·
Initiate patterns of gene expression
There are various families of transcription factors:
Homeodomain - Hox gene family
Zinc finger – a family of steroid-binding transcrition factors
Basic helix-loop-helix protein – myogenic regulatory
factors
Winged helix – hepatocyte nuclear factor-3
2. Signalling molecules. These are molecules that:
·
Exert their effect on other cells
·
Mediate most interactions e.g. induction
·
Bind to trans-membrane receptor molecules
·
Are mainly growth factors
Examples are:
Transforming growth factor-B (TGF-B) –
activate posterior Hox genes
Fibroblast growth factor (FGF) – activate
anrterior Hox genes
Nerve growth factor (NGF) –
stimulate growth of axons
Hedgehog proteins – mediate early
inductive interactions
3. Cell adhesion
molecules.
·
They are responsible for specific cell aggregation and sorting
·
Some are calcium-dependent
(Cadherins)
·
Some are calcium-independent (CAM)
An example is the
differentiation of the epiblast into neurectoderm (induced by the notochord)
and skin ectoderm by the differential expression of N-CAM and L-CAM in the
neurectoderm and skin ectoderm respectively.
***************************