Nerve Endings
Professor Alfred Cuschieri
Department of Anatomy, University of Malta.
Objectives
On completion of this unit the student should be able to:
o
Explain the significance of synapses in the nervous system
o
Name the structural and molecular characteristics of the pre- and
post-synaptic membranes, and the synaptic cleft.
o
Distinguish between different types of synapses and their functional
significance
o
Explain the structural and molecular characteristics of motor end plates
o
Name the different types of sensory nerve endings
o
Distinguish between stimuli and sensations
o
Test clinically the different modalites of sensation
o
Explain the structure and functions of muscle spindles
o
Name the different components of tendon, withdrawl and stretch reflexes
o
Distinguish the various modes of action of drugs acting on neuromuscular
junctions
o
Name the features of some common diseases resulting from defects of
neuromuscular transmission
Three main types of nerve endings
There are three main types of nerve endings:
1. Synapses – on other neurons
2. Motor nerve endings – on skeletal muscle
3. Sensory nerve endings – on receptors
Synapses
Synapses are sites of communication between neurons. At synapses the plasma membranes of adjacent cells are in close proximity and communication occurs by release of chemical neurotransmitters. Elsewhere in the central nervous system neurons are separated from one another by means of astrocytes (protoplasmic astrocytes in the grey matter and fibrous astrocytes in the white matter).
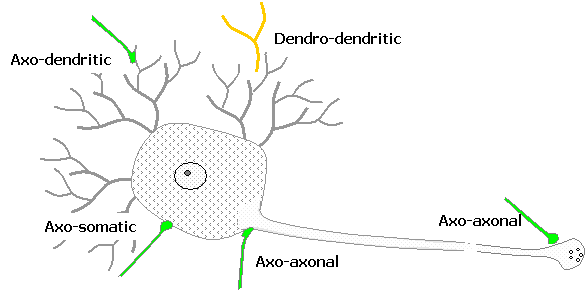
The most common type of synapse is axo-dendritric, but several other types of synapse occur. The axon usually has a dilated terminal expansion (bouton).
Synapses are unidirectional and consist of the following:
1. The axon terminal contains:
a. Neurotransmitter vesicles
b. The pre-synaptic membrane. This has electrically gated Ca 2+ channels that allow the influx of Ca 2+, when triggered by an electrical impulse. The increased Ca 2+ concentration stimulates the release of the neurotransmitter vesicles into the synaptic cleft by exocytosis.
c. Electron-dense cytoplasm, usually in patches, adjacent to the plasma membrane.
d. Mitochondria and microtubules
e. Coated vesicles and occasional lysosomes. These provide a system for the uptake of neurotransmitter from the synaptic cleft and its re-utilisation. This uptake system by pinocytosis also serves to remove the excess membrane generated by release of neurotransmitter vesicles.
2. The synaptic cleft:
a. Is a 20nm extracellular gap
b. Contains acid mucopolysaccharides
c. Contains an enzyme for inactivating the neurotransmitter
3. The post-synaptic membrane:
a. Is associated with electron-dense cytoplasm forming a sub-synaptic web.
b. Has specific receptors for the neurotransmitter
c. Has chemically-gated ion channels that are stimulated by the receptor-transmitter.
Note that:
Synapses may be stimulatory or inhibitory:
a. Some neurotransmitters are stimulatory e.g. acetylcholine (nicotinic) and L-glutamine, and the post-synaptic membrane contains chemically gated Na+ channels that produce depolarisation
b. Some neurotransmitters are inhibitory e.g. Gamma amino butyric acid (GABA), and the post-synaptic membrane contains chemically gated Cl- channels that produce hyperpolarisation
c. Some are modulators e.g. serotonin, histamine and adenosine, which modify the post-synaptic membrane potential via secondary messengers such as G-protein, which open or close K+ or Ca 2+ channels.
One neuron generally produces only one neurotransmitter that is used at all its synapses. Paradoxically, however, a neuron that produces a stimulatory neurotransmitter may also be inhibitory by acting via small internuncial neurons. Anterior horn cells are stimulatory to muscle and use acteylcholine, and also give collaterals that stimulate a Renshaw neuron, which in turn inhibits the anterior horn cells.
There are several known neurotransmitters. Examples are epinephrine, nor-epinephrine, dopamine, glycine, serotonin, GABA, enkephalins, substance P
And glutamic acid.
Synapses may occur between any parts of neurons:
Axo-dendritic synapses occur between an axon
termouton and a dendrite. At some sites
e.g. in cerebellum, the synapses occur on cytoplasmic projections known as
dendritic spines.
Axo-somatic synapses are between an axon and the
neuronal cell body. The neuronal cell
body and its dendritic tree form the receptive area of the neuron. Each synapse produces localised
depolarisation or hyper polarisation of the neurilemma. When the summated effects produce sufficient
depolarisation, the axon generates an impulse at the axon hillock or initial
segment of the neuron.
Axo-axonal synapses are between an axon terminal
and another axon, at one of the following critical sites: (a) an axon terminal
ending on the axon hillock, where it is critically located to inhibit or
facilitate the generation of an impulse; an axon terminal ending on another
axon terminal where it is critically situated to inhibit the release of
neurotransmitter.
Dendro-dendritic synapses occur between two dendrites
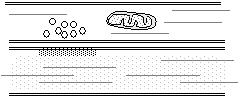
Synapses en passant do not involve a nerve
terminal, and may occur between any regions of two neurons, but are usually
dendro-dendritic. Such synapses influence the state of polarisation of the
adjacent neuron.
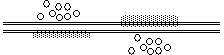 Reciprocal
synapses may occur between any regions of two neurons. These are in fact two adjacent synapses that influence one
another.
Reciprocal
synapses may occur between any regions of two neurons. These are in fact two adjacent synapses that influence one
another.
Neuromuscular junctions
Motor end plates are axon terminals
that end on skeletal muscle. They are very similar to cholinergic pre-synaptic
terminals. The
The axon terminal is devoid of a myelin sheath, and is thrown into terminal folds that contain:
o Neurotransmitter vesicles containing acetylcholine
o The pre-synaptic membrane, with electrically gated Ca 2+
o Mitochondria and microtubules
o Coated vesicles for the re-utilisation of neurotransmitters..
The synaptic cleft is a 20nm extracellular gap that contains acetyl cholinesterase for inactivating the acetylcholine neurotransmitter
The sole plate is a specialised region of the sarcolemma that is thrown into several junctional folds, but is otherwise basically similar to the post-synaptic membrane, having specific acetylcholine receptors coupled to chemically gated ion channels.
Sensory Nerve Endings
Sensory nerve endings may be classified functionally or morphologically.
Functional
classification
It is important to distinguish between a stimulus and the sensation it elicits. A nerve ending perceives a stimulus and generates a nerve impulse. It is perceived as a sensation by the brain. Sensations are classified according to their modality but there is no direct correlation with the morphology of nerve endings.
The following are different modalities of sensation.
o
Pain is the most primitive of sensations that results from noxious
stimuli. It may be tested by sensation to pin-prick
o Temperature may be tested by means of two test-tubes filled with warm and cold water. Extremes of heat and cold are interpreted as painful stimuli and may cause tissue damage.
o Touch (crude touch or fine touch) results from a tactile stimulus and gives rise to the awareness that one is being touched and the location of the stimulus. It may be tested using a blunt object.
o Discriminatory touch is the ability to discriminate between two points of simultaneous contact. It may be tested by the two-pin discrimination test, which determines the minimum distance at which one can distinguish two pins touching the skin simultaneously.
o Stereognosis is a special type of discriminatory touch that enables the distinction between different types of objects, including the discrimination of the texture and shape of the object. It is tested by asking the patient to identify a common object, such as a key, only by handling it.
o Vibration: the feeling of repeated stimuli caused by a vibrating object. This is a variant of discriminatory touch. It is tested using a tuning fork applied to a bony prominence.
o Proprioception is the transmission of information relating to the position of joints, the state of tension in ligaments and tendons, and the state of contraction of muscles. The most familiar form of proprioception is position sense in which the patient is aware of the postion of any part of his or her body without looking.
Morphological classification
Nerve endings are classified according to their structure. Nerve endings are always in contact with a transducer cell that converts a mechanical, thermal or chemical stimulus to an electrical potential in the nerve ending.
Free nerve endings are the branched terminations of the axons. Myelinated nerves lose their myelin sheath, and end in a number of branches that penetrate the area being innervated. They ramify between cells of the epidermis and form a plexus around hair follicles. The axon terminals end in small expansions (Merkel’s discs) that are in contact with specialised epithelial cells (Merkel’s cells or tactile domes). They are very sensitive to pain, temperature and crude touch. Note, for example that hairs are very sensitive to light touch and pain.
Free nerve endings are also common in somatic tissues including bones, joints and muscle.
Encapsulated nerve endings consist of branched axon enclosed in a discrete connective tissue capsule. There are several distinct varieties that are found in specific locations and subserve specific functions. Although several varieties have been described the following are well defined:
Meissner’s corpuscles
o Are found in thick skin of the palms and soles, and in the skin of the nipples and genitalia.
o Are sensitive to discriminatory touch.
o Are critically located in the dermal papillae where the overlying epidermis is thinnest
o Are oval structures
o Have branched, unmyelinated nerve terminals within their core
o Have transversely arranged, flat cells around the nerve terminals, considered to be modified Schwann cells of Neural crest origin.
o Are enclosed within an external connective tissue sheath
Pacinian corpuscles
o Are found in the deep tissues, particularly in the deep layers of the dermis, subcutaneous tissue, around joints, in the parietal pleura and peritoneum
o Are sensitive to deep touch, pressure and vibration
o Are large structures measuring 1-2 mm in diameter
o Have branched, unmyelinated nerve terminals in the core of the corpuscle
o Have concentric lamellae of flattened cells arranged longitudinally along the corpuscle
o Are enclosed within an external connective tissue sheath
Ruffini nerve endings
o Are stretch receptors
o Are located in the dermis of hairy skin
o Have a core of nerve endings and collagen bundles
o Are surrounded by a cellular capsule
Golgi tendon organs
o Are situated at musculo-tendinous junctions.
o Are enclosed within a capsule
o Have a core of intracapsular tendon bundles
o Have elaborately branched nerve endings twisted around the tendon core
o Have skeletal muscle fibres inserted into them
o Signal the strength of muscle contraction
o Are involved in reflexes that inhibit the homonymous (agonist) muscle and stimulate antagonistic muscles
There are several joint receptors located in the capsule and ligaments of joints. They are similar to Pacinian and Ruffini endings.
Neuromuscular spindles
Neuromusclar spindles are abundant in antigravity muscles (e.g. erector spinae and calf muscles), muscles of the neck and intrinsic muscles of the hand. They
consist of a group of 6 to 12 intrafusal muscle fibres, surrounded by connective tissue continuous with the perimysium.
The intrafusal muscle fibres are modified skeletal muscle fibres that have the characteristic striations but do not have their nuclei situated below the sarcolemma. Morphologically they are of two types:
1) Nuclear bag fibres contain a cluster of nuclei at the centre of the muscle.
2) Nuclear chain fibres have their nuclei arranged as a longitudinal chain at the centre of muscles.
Intrafusal muscles have a motor supply from the gamma efferents, ending in small motor nerve plates. Their sensory supply is of two types that differ in the arrangement of their endings:
(a) Anulospiral endings are arranged circularly around the centre of the fibre. They are most commonly found on nuclear bag fibres.
(b) Flower spray endings are complexly branching endings situated towards the ends of the fibres. They are most commonly found on the nuclear bag fibres.
Neuromuscular spindles are stretch receptors, sensitive to the state of tension in the muscle while it is actively contracting (active stretch) or when the relaxed muscle is passively stretched (passive stretch). They are responsible for mediating the so called ‘tendon’ reflexes such as the knee jerk, ankle jerk and biceps reflexes.
Nerve endings on smooth
muscle, cardiac muscle and secretory cells.
o These are the nerve endings of postganglionic autonomic nerves.
o They are always free nerve endings and do not have motor end plates or other specialised endings
o They may be cholinergic or adrenergic
o Gastrointestinal smooth muscle and cardiac muscle contract independently of motor nerve stimulation. The autonomic endings on these two types of muscle may affect the rate of contraction of the muscles.
Reflexes are simple nerve circuits that involve
o A receptor ending
o One or more synapses
o An effector ending
In a tendon reflex, e.g. the knee jerk:
o The receptor endings are the neuromuscular spindles (e.g. in the quadriceps muscle) that perceive passive stretch
o The sensory neuron terminates on an internuncial neuron in the posterior horn of the spinal cord, which in turn synapses with an anterior horn cell supplying the homonymous (same) muscle
o The axon of the anterior horn cell ends on a motor end plate that stimulates the muscle to contract
The withdrawl reflex is the withdrawl of a part of the body from a harmful stimulus, such as sudden and unexpected contact with a sharp or very hot object.
o The receptor is the free nerve ending in the skin
o Synapses occur between the sensory, internuncial and motor neurons in the spinal cord
o Stimulation of flexor muscle occurs at the motor end plates
The inhibitory stretch reflex is elicited by excessive tension on a tendon.
o The receptor is the Golgi tendon organ
o Synapses occur in the spinal cord with inhibitory internuncial neurons ending on the homonymous muscle and with stimulatory internuncial neurons on the antagonistic muscles.
o Stimulation of antagonistic muscles occurs at the motor end plates
Other more complex reflexes, such as the eye reflex and head-turning reflex will be dealt with elsewhere
Drugs That Act On Motor End Plates
Non-depolarising Neuromuscular blocking agents
This class of drugs includes substances that combine with, and block, the acetylcholine receptors, but do not depolarise them. They are also known as competitive inhibitors of acetylcholine. They include the aminosteroid group (pancuronium, rocuronium and vecuronium), and the benzoquinolinium group (atracurium,cisatracurium, mivacurium and gallamine). They are used for producing muscle relaxation in anaesthesia. Their effect wears off
Depolarising Neuromuscular
blocking agents
These substances, such as suxamethonium, are similar to acetylcholine and produce depolarisation of the motor end plate, but are not inactivated by acetylcholinesterase, so that their action persists. They produce a single muscle contraction, followed by muscle paralysis that persists until the depolarisation persists. They are also used for producing muscle relaxation under anaesthesia.
Anticholinesterases
These substances, such as physostigmine, neostigmine and edrophonium, inhibit the action of acetylcholinesterase, so that the action of acetylcholine is prolonged. They reverse the action of non-depolarising blocking agents by making more acetylcholine available for competing with the blockers. However, they prolong the action of depolarising blocking agents.
Diseases associated with
defects in neuromuscular transmission.
Myaesthenia gravis is an autoimmune disorder in which antibodies are produced against acetylcholine receptors. There is consequently a progressive muscle weakness characterised by general muscle weakness, ptosis (drooping of the eyelids), diplopia (double vision due to paralysis of the ocular muscles), an expressionless face (paralysis of facial muscles), dysphagia (difficulty in swallowing, due to paralysis of pharyngeal muscles) and dysarthria (difficulty in talking).
Hypokalaemic periodic paralysis is caused by low potassium levels in the blood. It causes defective functionng of the voltage-gatedcalcium channels, producing sustained depolarisation of the postsynaptic membrane and consequent paralysis.
Organophosphorous poisining is caused by various substances including various insecticides such as malthion. It produces irreversible inactivation of acetylcholinesterase and consequent prolonged depolarisation of the postsynaptic membrane. Atropine protects receptors from high acetylcholine levels and is used for treatment.
Snake venom poisoning blocks acetylcholine receptors at neuromuscular junctions, causing muscle paralysis. Antivenons are available.
![]()
