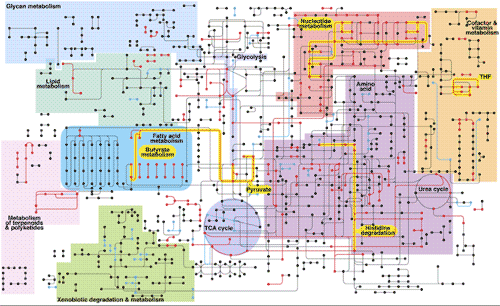
Cell metabolism is based on a network of organic reactions in water, each catalyzed by an enzyme. Water has the advantage that it is a good solvent, and is abundant on Earth.
Energy
Adenine
Ribose
Tri/Di-Phosphate
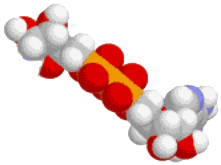
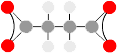
ATP⇆ADP
Cells require an energetic molecule to drive some of the unlikely metabolic reactions. It should be stable enough not to react needlessly on its own, yet not so stable that it doesn't release its energy readily, hence slowing down the reactions. The molecule that is used is ATP. It can transfer a phosphate to a molecule with about 0.31eV of energy, priming it for a reaction. Each individual ATP molecule is used and recycled about every .
On Earth, there are two ready sources of energy — light from the sun, and the outgassing of reduced compounds such as hydrogen sulphide and hydrogen into the oxidized atmosphere.
Reducer
Nicotinamide Adenine Dinucleotide H ⇆ NAD+
Oxidizing reactions occur spontaneously in Earth's oxygen-rich atmosphere; all naturally occurring molecules are oxidized (e.g. CO2), so to utilise them, cells require a strongly reducing molecule. The molecule used is NADH (or its variant NADPH), which readily gives a hydrogen atom and an electron.
How are ATP and NADH generated?
Cells generate an electric potential of about across their membranes by pumping protons from inside to outside of the membrane, and use the protein concentration imbalance as a ready source of energy (like a battery). Details vary depending on the species.
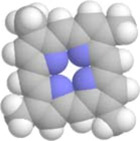
A protein complex, called ATP synthase, allows protons to pass back to the negatively charged interior, in the process causing its internal parts in the upper portion to rotate. The lower part, connected to it, rotates in unison and forces ADP to accept another phophate ion and become ATP, ready to be used by the metabolic reactions.
p+
3D working model of ATP Synthase
But how are protons pumped in the first place?
Chemotrophic cells are rich in NADH; consumers (heterotrophs) generate it from ingested reduced organic molecules such as carbohydrates () and fats () that they consume. They do so by using NAD+ to oxidize them to CO2 and H2O.
Such cells react NADH with an oxidizing molecule, ultimately to pump proteins. The protein Complex I below (or a variant Complex II) has a sequence of polypeptides (flavoproteins) and Fe-S centers with matching energy levels that transfer protons to the outside and electrons to a quinone molecule, called coenzyme Q, attached to it. This reduced quinone is then released into the membrane.
The net effect of Complex I is that NADH is oxidized to NAD+, coQ is reduced to coQH2 and up to four protons are transported across the membrane.
p+
e-
p+
A second protein complex Complex III accepts the diffusing reduced coenzyme Q, and transfers its protons to the outside and the electrons to a cytochrome molecule attached to it. The net effect is that coQ is oxidized back to Q, cytochrome is reduced, and four protons are transported across the membrane.
e-
p+
e-
p+
The reduced cytochrome molecule diffuses away in the water, held to the positive protons around it...... until it meets protein Complex IV which takes its electron and passes it along a cascade of oxidizing agents inside it, to an oxidizing molecule (e.g. O2) inside the cell.
e-
p+
p+
The net effect of all these reactions is that protons are pumped across the membrane, while NADH is oxidized to NAD+ and an oxidising agent (oxygen O2) is reduced (to water H2O).
Phototrophic cells use a very similar system, but since they are not rich in NADH, they produce reduced quinone by oxidizing some other molecule, most commonly water, using light energy. Instead of the first protein complex, they have a photo-system consisting of an array of chlorophyll and carotenoid molecules which capture light and transfer its energy to a central chlorophyll molecule. There, an excited electron is transferred to an attached quinone molecule which then detaches. Its electron is replenished by water (usually), which is turned to oxygen.
Other producers (autotrophs) generate NADH from the reduced quinone in the membrane, using a membrane protein and either light (phototrophs) or ATP as an energy source (essentially protein complex I run in reverse).
e-
p+
p+
e-
e-
p+
Additionally, phototrophs have membrane proteins similar to Complex III; but Complex IV is replaced by a protein that harvest light energy to transfer electrons from reduced cytochrome to NAD+, thus producing NADH.
The net effect, for phototrophs, is that protons are pumped across the membrane, while NAD+ is reduced to NADH and a molecule (water H2O) is oxidised (to oxygen O2).
Overall, the net reactions performed by cells to generate energy and synthesize organic molecules are, for example:
Autotroph
Heterotroph
Phototroph
light + H2O +CO2 → O2 + CH2O
light + H2S + CH2O → S + H2 + CH2O
Chemotroph
4H2S + O2 +CO2 → 4S + 3H2O + CH2O
CH2O + O2 → CO2 + H2O
(CH2O stands for carbohydrates in general.)
Core Metabolic Reactions
The cell needs to keep a balanced supply of 'building blocks'. These are the set of 3-carbon molecules shown below, from Glycerol and Glyceraldehyde (G3P) to Pyruvate.
When the cell is rich in ATP and NADH, the balance shifts to the upper reduced compounds; otherwise it shifts to the lower set of oxidized compounds.
GlycerolP
Lactate
G3P (GlyceraldehydeP)
DHAP
Pyruvate
PEP
PGA (GlycerateP)
A number of enzymes convert between the molecules, keeping them in balance (grey arrows). For example, enolase keeps PEP in balance with PGA (glycerate). Other enzymes, called dehydrogenases, keep balance between upper and lower sets of molecules (red arrows), e.g. DHAP⇆PGA. They require NADH to move up (reduce) and NAD+ to move down (oxidize or dehydrogenize).
Small neutral molecules, such as these, are phosphorylated to prevent them from diffusing across the membrane; the phosphate is transitory and not an essential biological part of the molecule. Very small molecules such as oxygen, acetate, formate and methylene are carried by or joined to larger molecules, called coenzymes, to prevent them from diffusing away and to slow them down enough for enzymes to handle them.
The Citrate Cycle
The core reactions of the cell convert pyruvate into larger molecules that are the base for synthesizing amino acids, etc. For heterotrophs (consumers) this cycle breaks down pyruvate into three CO2 and is their main way of generating NADH.
Citrate
Isocitrate
Ketoglutarate
Succinate
Fumarate
Malate
Oxaloacetate
Pyruvate
Acetate
First, pyruvate is oxidised to acetate, giving off CO2.
Another enzyme, citrate synthase then combines acetate with oxaloacetate to form the more reduced citrate. Using a sequence of dehydrogenases, citrate is broken back down to oxaloacetate, while giving off two CO2 and four NADH.
pyruvate
acetate
oxaloacetate
malate
fumarate
succinate
ketoglutarate
isocitrate
citrate
+NADH
If the amount of oxaloacetate falls too low, a carboxylase enzyme combines pyruvate with HCO3- to form oxaloacetate. Conversely, if there is too much, another enzyme converts it to PEP (and hence to pyruvate).
Autotrophs do not want to give out CO2, so isocitrate is not oxidised to ketoglutarate but directly to succinate; effectively, 2 acetate molecules are converted to malate:
isocitrate
succinate
malate
acetate
glyoxylate
Synthesis/Glycolysis of Small Carbohydrates
The other important molecules are sugars, with 3 to 7 carbons:
G3P
E4P Erythrose
R5P Ribose
G6P Glucose (≅F6P fructose)
S7P Sedoheptulose
Each of these phosphorylated sugars has several isomers: placement of OH (e.g. glucose/galactose), linear or cyclic forms, placement of double-bonded O (e.g. glucose/fructose, ribose/ribulose). Enzymes called isomerases convert between these forms.
Other enzymes, called aldoses, bond together or break up these molecules, or break and reassemble them: , , ,
Thus all these simple sugars are kept in balance by ten enzymes; an excess of G3P results in an increase in the other sugars, or vice versa. The gateways to these reactions are controlled by enzymes called kinases which add or remove phosphate groups, thus regulating whether the sugars are primed to react, e.g., F6P and S7P to F6PP and S7PP, and whether PEP is converted to pyruvate.
Autotrophs have several more enzymes that synthesize simple molecules. Two of the most important are:
Rubisco captures CO2 using Mg2+, adds it to R5P to form two PGAs; although this is a slow reaction, it is the main way that autotrophs generate 3-carbons.
Several enzymes working together convert E4P + PEP to chorismate, a ringed molecule.
Control the metabolism by switching on/off some enzymes:
Acetate
Pyruvate
G3P
Oxalate
Malate
Succinate
E4P
Ketoglutarate
RiboseP
Citrate
GlucoseP
S7P
Biosynthesis of Amino Acids
The building blocks are used to create amino acids, nucleic acids, and lipids, that in turn are built up into proteins, DNA, and membrane of the cell.
The simple amino acids are built up from the common cellular molecules by transamination enzymes: the exchange of an O or OH atoms by NH2:
Pyruvate
Alanine
Pyruvate
Alanine
PGA
Serine
Glycine
(by removal of OH)
Oxaloacetate
Aspartate
Asparagine
Ketoglutarate
Glutamate
Glutamine
The transamination that produces glutamine (and glutamate) is reversible; in fact glutamine is mostly used to store ammonia as a source to transaminate other molecules.
The other amino acids require more specialized pathways:
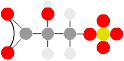
Replacement of OH with SH (or even SeH)
Serine
Cysteine (or seleno-cysteine)
Glutamate
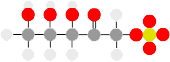
Two pyruvate molecules combine to form acetolactate , then
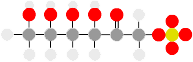
Aspartate is reduced to , then to
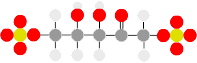
Chorismate is the source of ringed amino acids:
On average, protein synthesis uses up most (3/4) of the cell's energy.
Biosynthesis of Nucleic Acids
The starting point is aspartate, which when reduced with carbamoyl, gives the basic pyrimidine cyclic molecule:
Combining pyrimidine with
ribose gives uracil and thymine
ribose, followed by transamination, gives cytosine
nicotine, gives NAD
Enzymes that transaminate ribose (R5P), then combined with glycine, formate, CO2, aspartate, and a second formate, added in succession, produce the basic inosine purine molecule.
Inosine, transaminated, is essentially Guanosine; transaminating the extremal oxygen gives Adenosine. Further work produces histidine, the most difficult amino acid to synthesize.
Biosynthesis of Lipids
Joining acetyl-coA together one by one, and removing the oxygens gives the lipid molecules.
This process can run in reverse to decompose lipids into acetate.
Joining the acetyl-coA in a different way, and reducing it, gives isoprene. When repeated, it becomes a quinone molecule.
Succinate combined with glycine gives aminolevulinate which pair up and join as porphobilinogen...
...four of which combine to form the basic Porphyrin found in cytochromes.
Metabolic Control
The network of reactions is controlled by the presence of specific enzymes.
Each of these enzymes can be switched on/off (activated/inhibited) by a particular combination of effector or repressor molecules that bind to them in regions that affect the active site; in addition, some can be permanently switched off by being cleaved. So as one product increases in concentration, it may switch on/off other enzymes, setting off other changes along the network.
There are two main overriding states of the cell:
Well-fed state: switch on biosynthesis, with generation of glucose, lipids, and amino acids. Excess glucose is stored as a polysaccharide, e.g. glycogen, starch. Too high a level of amino acids leads them to be catabolized.
Starved state: switch on glycolysis; if too low in glucose, then lipids are oxidized; if still too low, then amino acids are catabolized. Cells may secrete enzymes externally to lyse (digest) polysaccharides, lipids and proteins.
However, in only a very few cases is the complete network and regulation of the cell metabolism known. There are simply too many sub-states and sub-networks. For example, most have low/high oxygen states, a 'circadian' clock network that anticipates day/night for optimal control, etc.
The cellular metabolism is quite resilient to "shocks" in concentrations; if some molecule is being depleted, there is usually some other pathway that can regenerate it (as long as ATP and NADH values themselves aren't low). However, because of its complex feedbacks, faulty proteins can easily disrupt the balance of metabolites. To prevent this, the inside of a cell is continually purified — marked proteins are broken down by special proteins into short sequences of amino acids, while ketones are metabolized; small molecules are not allowed to concentrate, but are slowly removed by adding OH to make them soluble enough to diffuse away; small molecules in the membrane are captured by special glycoproteins.
The reactions need certain optimum conditions to work properly:
water : solution cannot be too concentrated, i.e., saline, or dry;
: to achieve this, the inside of the cell is buffered by K+ and H2PO4-;
temperature : usually about tolerance about some optimum (, depending on species); cannot withstand boiling and freezing of internal water; macromolecules are denatured outside the tolerated temperature;
radicals (O2-, OH, NH3, CO, CN, H2S, ...) poison (react with) macromolecules; cytochrome enzymes oxidise unusual small molecules to render them inactive and soluble so they can diffuse out of the cell.
The effective metabolic efficiency is of the order of 20-25%. Note that, at normal temperatures (20-30C), small molecules (e.g. O2) take to diffuse , medium molecules (e.g. ATP) take , and large ones (proteins) take .
Heterotrophs
Many consumers do not need to synthesize every compound they need, especially those that require a lot of energy to produce. Instead they acquire them from their organic intakes, which then become essential nutrients: some amino acids (those that are not produced by a simple transamination), purines and pyrimidines, vitamins (for coenzymes, e.g. NADH = niacin B3, FADH = riboflavin B2, coA = B5).
Rather consumers are often faced with excess amino acids: these are deaminated (NH4+ removed) or transaminated (NH3 replaced by O) to leave ketones that are oxidized in a similar way to the lipids; the ammonia has to be removed as waste.















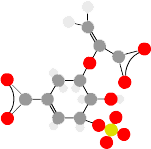
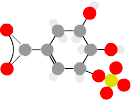
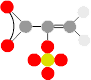


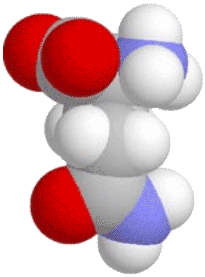
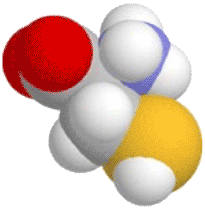
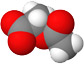 , then
, then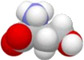 , then to
, then to



 . When repeated, it becomes a
. When repeated, it becomes a  which pair up and join as porphobilinogen...
which pair up and join as porphobilinogen...