6
CHROMOSOMES
IN MITOSIS AND
MEIOSIS
Professor A. Cuschieri
Department of Anatomy
University of Malta
OBJECTIVES:
By
the end of this session the student should be able to:
* Explain how DNA is organised in chromosomes
* Distinguish between mitosis and meiosis
* State where mitosis and meiosis occur in the human
* List the events that occur during mitosis and
meiosis
* Explain the
significance of chromosome recombination
* Name the main features
by which chromosomes are identified
* Give examples of aneuploidies and explain how they arise
* Classify main types of chromosome
abnormalities
* Distinguish between balanced and unbalanced
chromosome rearrangements
* Deduce how unbalanced chromosome
anomalies may occur in the offspring of carriers of balanced rearrangements
INTRODUCTION
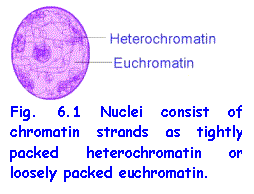 In living cells DNA is always associated with proteins and is organised
to form chromatin strands and chromosomes.
In interphase nuclei, i.e. nuclei of non-dividing cells, there are 46 extremely long chromatin
strands, one for each chromosome. In
places, the strands are loosely arranged to form euchromatin and in others they are densely
packed heterochromatin. Everyone knows how thread becomes hopelessly
tangled unless it is kept neatly organised on reels or balls. It is most
remarkable how these extremely long, thin threads of chromatin do not become
tangled and can sort themselves out and become neatly wound to form the
chromosomes. Chromosomes are neat ways
of packaging the DNA during mitosis and meiosis for easy distribution to
daughter cells.
In living cells DNA is always associated with proteins and is organised
to form chromatin strands and chromosomes.
In interphase nuclei, i.e. nuclei of non-dividing cells, there are 46 extremely long chromatin
strands, one for each chromosome. In
places, the strands are loosely arranged to form euchromatin and in others they are densely
packed heterochromatin. Everyone knows how thread becomes hopelessly
tangled unless it is kept neatly organised on reels or balls. It is most
remarkable how these extremely long, thin threads of chromatin do not become
tangled and can sort themselves out and become neatly wound to form the
chromosomes. Chromosomes are neat ways
of packaging the DNA during mitosis and meiosis for easy distribution to
daughter cells.
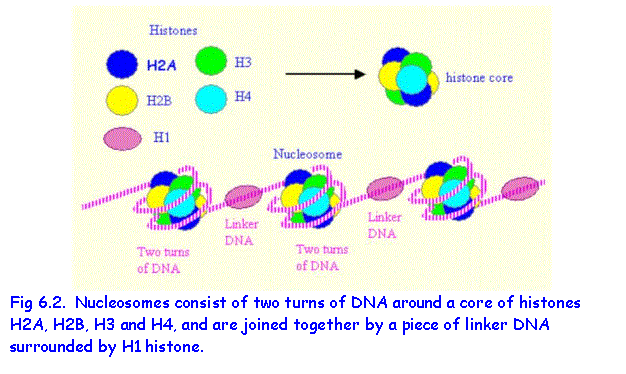
NUCLEOSOMES
The DNA in nuclei and chromosomes is associated
with histone proteins to form chromatin strands (Fig.
6.1). Eight histone protein molecules,
consisting of four pairs of histones H2A, H2B, H3 and H4 are organised into a histone core. A length of DNA consisting of about 140 base
pairs makes two turns around the histone core thus forming a nucleosome. The DNA extending between two nucleosomes is
called linker DNA and
is associated with histone 1.
 CHROMATIN STRANDS
CHROMATIN STRANDS
The beaded chain of nucleosomes is wound into a
spiral containing six nucleosomes per turn. H1 histone is responsible for the secondary coiling. The coiled coil filaments are the chromatin strands. These are further folded in complex
ways to form tightly packed chromosomes
in dividing cells or loosely packed chromatin in interphase nuclei. Non-histone chromosome proteins regulate the
unfolding of the chromatin so that transcription can take place.
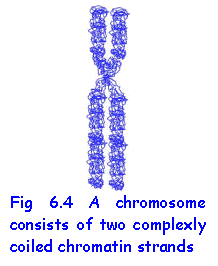 CHROMOSOMES
CHROMOSOMES
Chromosomes become evident as discrete
structures only during cell division, whether mitosis or meiosis. In both processes the chromatin strands
replicate to form two identical strands and then coil up or condense to form
chromosomes. Each chromosome is thus a duplicate structure consisting of two chromatids attached to one another at the centromere.
MITOSIS AND THE CELL CYCLE
Mitosis is the process of cell division in
which two identical daughter cells are formed. The cells replicate the genetic
material so as to make two identical copies of DNA, which are then distributed
to the daughter cells. In a population
of dividing cells the changes undergone by a cell are referred to as the cell cycle (Fig. 6.5).
 Each cell cycle starts with a newly
formed daughter cell at the end of a mitotic division and involves the
following stages:
Each cell cycle starts with a newly
formed daughter cell at the end of a mitotic division and involves the
following stages:
1. Gap 1 phase (G1) during which the cell performs its
normal functions
2. Synthesis
phase (S) during which replication (synthesis) of DNA occurs.
3. Gap 2
phase (G2) during which
the cell cytoplasm increases in bulk and forms new organelles in preparation
for cell division
4. Mitosis (M), the actual process of cell division, which
consists of a number of stages:
a.
prophase: the
chromosomes condense and become distinct
b.
metaphase: the
chromosomes collect at the equator of the cell and a mitotic spindle is
formed. The spindle is formed of
microtubules which attach to the centromeres of the chromosomes
c. anaphase: the chromosomes split at the centromere and
the two chromatids move to opposite poles of the cell
d. telophase: the chromosomes at opposite poles
de-condenseand become enclosed in a nuclear envelope forming the two daughter
nuclei.
Mitosis occurs in all cells during embryonic
and foetal development. In postnatal
life it occurs in growing tissues. In
adults it is restricted to certain sites, namely:
* bone
marrow for haemopoiesis;
*
epidermis of skin, nails and hairs;
*
epithelium lining the intestinal tract;
*
connective tissue and bone during repair following injury.
In
other tissues e.g. muscle, liver and most internal organs mitosis occurs at a
very slow rate. In some tissues, notably the central nervous system, the cells
lose their ability to divide in postnatal life.
MEIOSIS
Meiosis occurs only in the germ cells i.e. the
cells that form the spermatozoa in males and the oocytes (ova) in females.
Whereas somatic cells have a diploid
number of chromosomes, i.e. the chromosomes are in pairs, the spermatozoa and
oocytes have a haploid
set of chromosomes, i.e. only one of each pair of chromosomes. In man, the diploid number of chromosomes
is 46. The haploid set found in the
germ cells consists of 23 chromosomes.
Meiosis is the process of reduction division in which the chromosome
number is halved from diploid to haploid. This is necessary because
fertilisation involves the fusion of a male and a female gamete to form a
zygote thus restoring the normal chromosome number. However, meiosis serves a second extremely important function -
genetic recombination.
RECOMBINATION
 In all diploid cells, the chromosomes
occur in homologous pairs,
derived one from the mother and one from the father. During meiosis, homologous chromosome pairs associate with one
another and exchange segments of DNA. This is referred to as crossing over. When the
chromosomes separate, each chromosome is a mixture of the original maternal and
paternal chromosomes. This causes a
reshuffling or recombination
of genes. Recombination is an important means of ensuring genetic variation among the offspring. Crossing over involves breakage and re-union
of the chromatin strands.
In all diploid cells, the chromosomes
occur in homologous pairs,
derived one from the mother and one from the father. During meiosis, homologous chromosome pairs associate with one
another and exchange segments of DNA. This is referred to as crossing over. When the
chromosomes separate, each chromosome is a mixture of the original maternal and
paternal chromosomes. This causes a
reshuffling or recombination
of genes. Recombination is an important means of ensuring genetic variation among the offspring. Crossing over involves breakage and re-union
of the chromatin strands.
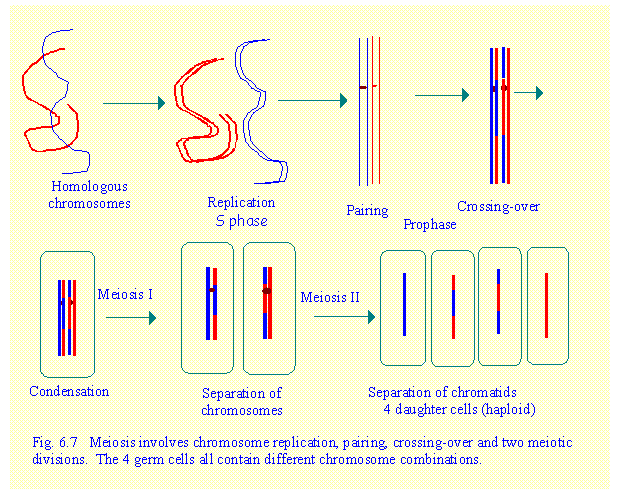
Cells undergoing meiosis pass through G1, S and
G2 phases. During S phase chromosome
replication occurs. The process of
meiosis consists of two consecutive
divisions known as Meiosis 1 and Meiosis 2.
a.
Meiosis 1. Recombination occurs during the prophase of
meiosis 1. Pairing and crossing-over of segments occurs between homologous chromosome
pairs. This is accompanied by condensation of chromosomes. Metaphase of meiosis 1 results in separation of homologous chromosomes. Note that this
is different from mitosis in which chromatids separate. The two daughter
cells contain a haploid set of chromosomes
(only one of each pair).
b.
Meiosis 2. This occurs immediately after
meiosis 1, when the chromosomes are still condensed. Therefore there is no prophase. Metaphase of meiosis 2 results in the separation of
chromatids. 4 germ cells are formed
each containing a haploid set of chromatin strands.
These stages are summarised in Fig 6.7
![]()
Note that there is no prophase preceding
meiosis 2. The chromosomes are already
duplicated and condensed at the end of meiosis 1 and so proceed immediately to
meiosis 2.
Remember:
*
Meiosis results in four germ cells each receiving a haploid set of
chromosomes
* The chromosomes in each germ cell are
homologous but not identical to one another.
* The germ cells contain one set of genes
* As the sites of crossing over are random, the
patterns of recombination of genes and chromosomes are almost infinite.
CHROMOSOME
ANALYSIS
Chromosomes are visible as discrete structures only during cell
division. Chromosome analysis requires
a source of dividing cells, usually obtained from blood lymphocytes, which have been stimulated to enter mitosis. Chromosomes are examined in cells in
metaphase.

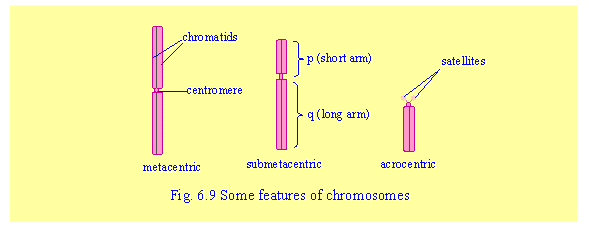
Chromosomes are classified as:
a.
Metacentric (centromere close to the centre) –
chromosomes 1, 3, 16, 19 and 20 are metacentric of different size.
b.
Submetacentric (centromere
some distance away from the centre).
c.
Acrocentric (centromere close to one end) –chromosomes 13, 14 and 15 (large
acrocentric) and chromosomes 21 and 22 (small acrocentrics). All the acrocentric chromosomes have a very
small short arm and a pair of "satellites" attached by a narrow stalk. The satellites
consist of repeated copies of the same genes that form r-RNA and are known as
the nucleolus organisers
because they associate together within the nucleus to form the nucleolus.
d.
Sex chromosomes
– the X and Y chromosomes are unequal in size.
Females have XX and males have
XY sex chromosomes.
Note that the short arm of a
chromosome is designated as "p"
and the long arm as "q".
CHROMOSOME ABNORMALITIES
These are classified into aneuploidy (numerical
abnormalities) and structural
chromosome abnormalities.
ANEUPLOIDY
Aneuploidy is an abnormality in chromosome
number and involves the loss or gain of chromosomes. The most common
aneuploidies are:
1. Trisomy
Trisomy affects one particular chromosome and
occurs if there are three instead of a pair of homologous chromosomes. The
karyotype has 47 chromosomes. Trisomy involves a large number of extra genes.
This causes congenital anomalies that are recognisable as characteristic clinical
syndromes. The most common examples
are:
Trisomy 21
- Down's Syndrome
occurs in about 1 in 600 births.
Trisomy 13
- Patau's Syndrome occurs in about 1 in 5,000 nirths
Trisomy 18
- Edward's Syndrome
occurs in about 1 in 8,000births
Trisomies
involving the sex chromosomes are much more frequent; they include XXY (Klinefelter Syndrome), XXX and XYY.
Aneuploidies involving other chromosomes are
rare as they are often incompatible with life and end as spontaneous
miscarriages.
2. Monosomy
Monosomy involves the loss of a
chromosome. The karyotype contains 45
chromosomes. Monosomy involves the loss
of a large number of genes. Most monosomies are incompatible with life and are
seen only in spontaneous abortions. The
example that occurs quite commonly and is compatible with life is monosomy X (Turner Syndrome).
3. Other aneuploidies
These include tetrasomies and pentasomies (4 or 5 homologous chromosomes
instead of a pair). They are very rare
and are found only involving the sex chromosomes.
Aneuploidy usually arises as a result of an
error in meiosis when a pair of homologous chromosomes fails to separate. The phenomenon is termed non-disjunction. It is often a defect of the mitotic spindle,
which fails to attach to one of the chromosomes. Non-disjunction may occur in either the maternal or the paternal
germ cells. Trisomy 21 arises more commonly in maternal than in paternal
meiosis. Increasing maternal age is a contributory factor in causing
non-disjunction.
4. Mosaicism
Mosaicism is the condition in which an
individual has cells with different chromosome complements. Mosaicism usually
results from non-disjunction during mitosis in the zygote. Consider a zygote
with a normal chromosome complement of 46.
If non-disjunction affecting one chromosome pair occurs in the first
mitotic division, one of the daughter cells would have 47 chromosomes and the
other 45. If non-disjunction occurs in the second or subsequent mitosis, there
will be three cell lines including the original 46, as well as 47 and 45
chromosomes. During subsequent mitosis
certain cell lines, especially monosomies are often eliminated or much
reduced.
Certain aneuploidies exist only in mosaic form
e.g. trisomy 8 , trisomy 9 and trisomy 22
5. Polyploidy
Normally, somatic cells are diploid containing
pairs of chromosomes. In polyploidy there are sets of three or four
chromosomes. In triploidy there are 23 x 3 = 69 chromosomes and in tetraploidy
there are 23 x 4 = 92 chromosomes. Triploidy is very rare and is invariably fatal
at or shortly after birth; it is found more commonly among abortuses and
stillbirths. Triploidy may results
from a complete failure of formation of the mitotic spindle or of cell division
during meiosis.
STRUCTURAL CHROMOSOME ANOMALIES
 The more common types are:
The more common types are:
1. Deletions
A single chromosome break occurs in one of the
chromosomes and the broken fragment is lost.
This results in the loss of a number of genes. Fig. 6.10 shows a deletion of the short arm
of chromosome 5 resulting in the cri-du-chat syndrome. The karyotype would be 46,XX 5p- or 46,XY
5p-.
2. Translocations
These are
usually classified into two types:
 a. Reciprocal translocations
a. Reciprocal translocations
Part of a chromosome breaks off and becomes
attached to another chromosome.
Usually, there is a reciprocal
translocation between two chromosomes where parts of two chromosomes break
off and become attached to the other chromosomes. Fig.6.11 illustrates a reciprocal
translocation between chromosomes 2 and 9.
Two abnormal translocation chromosomes are thus formed. The karyotype
would be 46,XX t(2;9).
Note that the broken end of a chromosome cannot reattach to the intact end of a chromosome, as this would be stable. It can only attach to another broken site. If there is only one break point, a rearrangement cannot be formed and the acentric fragment would be lost as occurs in a deletion.
b. Robertsonian translocations
A Robertsonian translocation occurs between two
acrocentric chromosomes, which appear to be attached to one another end to end.
Fig.6.12 shows a Robertsonian translocation between chromosomes 14 and 21.
However, this is not really an “end to end" translocation but a reciprocal
translocation in which the long arms of the two chromosomes fuse. The short
arms of both chromosomes also fuse to form a minute fragment, which is usually
lost. The loss of these short arms does
not cause any abnormal effects since they contain the nucleolus organisers of which there are many
other copies on the other acrocentric chromosomes. The karyotype would be 45,XX t(914;21) or 45,XY t(2;9).
c. Unbalanced translocations
Translocations, as described above, are balanced. They do not
involve any loss or gain of genes. Individuals who carry a balanced
translocation are phenotypically normal.
However, problems may arise during pairing of homologous chromosomes in
meiosis and may cause unbalanced
translocations in the offspring. The following examples illustrates how
unbalanced rearrangements may arise from balanced translocations in the
parents:
Example 1. Marika is a carrier of a
translocation between chromosomes 14 and 21.
What would be the possible
outcome for her offspring?
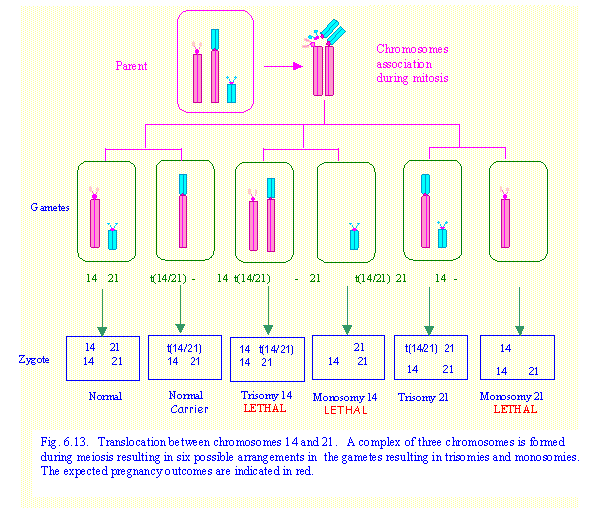
In this case Marika has 45
chromosomes. There is only one each of normal chromosomes 14 and 21 and a
translocation chromosome 14/21. Pairing
of homologous chromosomes during meiosis cannot occur in the normal fashion.
The three chromosomes associate together as shown in Fig. 6.13 so that there is
still point-to-point pairing of corresponding chromosome regions. When the three chromosomes separate, two
chromosomes go to one gamete while the remaining chromosome moves to the other
gamete. There are six possible ways in
which these chromosomes can be sorted as shown in Fig.6.13.
![]() The zygote receives a set of
chromosomes 14 and 21 from the normal parent.
The possible outcomes in the zygote are (a) normal – 46,XX or XY; (b) translocation carrier 45, XX or XY;
(c) trisomy 14 – karyotype 46,XX /XY,
-21 + t(14;21); (d) monosomy 14 –
karyotype 45,XX /XY, -14; (e) trisomy 21 – 46,XX/XY, -14 + t(14;21); (f) monosomy 21 – karyotype 45,XX /XY, -21.
The zygote receives a set of
chromosomes 14 and 21 from the normal parent.
The possible outcomes in the zygote are (a) normal – 46,XX or XY; (b) translocation carrier 45, XX or XY;
(c) trisomy 14 – karyotype 46,XX /XY,
-21 + t(14;21); (d) monosomy 14 –
karyotype 45,XX /XY, -14; (e) trisomy 21 – 46,XX/XY, -14 + t(14;21); (f) monosomy 21 – karyotype 45,XX /XY, -21.
Monosomy 14, monosomy 21 and most cases of
trisomy 14 would be lethal in early pregnancy and would result in spontaneous
abortion. Therefore, 1 out of 3 babies
born at term would be expected to have translocation trisomy 21 (translocation
Down Syndrome), 1 out of 3 would be a translocation carrier and 1 out of 3 will
be normal. These are theoretical risks. In practice it is found that the actual or empiric risk for a carrier
parent having a baby with Down syndrome is less than 1 in 3. The reason for this is not understood.
Example 2: Raymond has a reciprocal translocation between the short arms of
chromosomes 2 and 9. What are his
chances of having a normal child?
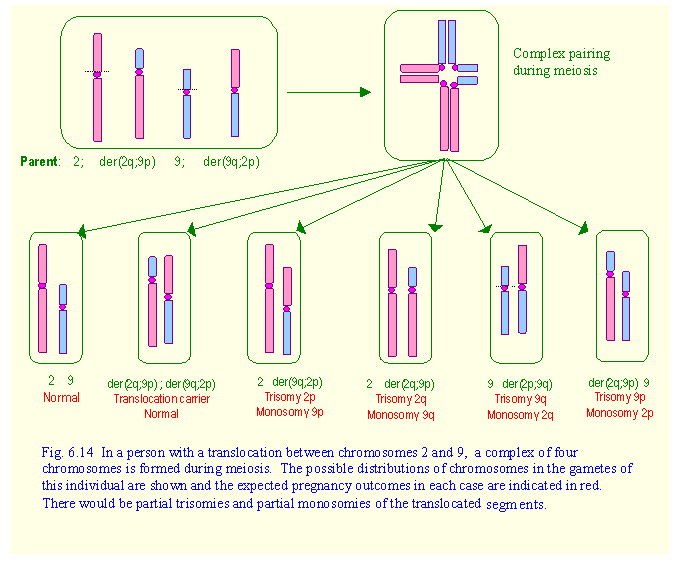
In this case, the short arm of
chromosomes 2 (2p) has been translocated to the long arm of chromosome 9 (9q),
while the short arm of chromosome 9 (9p) has been translocated to the long arm
of chromosome 2 (2q). This gave rise to
two derivative chromosomes, which are designated in a simplified way as
der(2q;9p) and der(9q;2p). Here again
chromosome pairing during meiosis will be very complex. In fact, the four chromosomes associate
together in a cross-like manner so that there is point to point correspondence
of chromosome and crossing over is still possible (Fig.6.14).
When the four chromosomes separate, two will go
to one gamete while the other two go to the other gamete. This gives rise to six possible combinations
as shown in Fig.6.14. The outcomes would be partial trisomies and partial
monosomies of the translocated segments of chromosomes 2 and 9. All of these
would be incompatible with life. Two out of three pregnancies would end in
spontaneous abortions, while 1 in 3 would result in normal children, or ones
who are carriers like their parent. Such individual are often discovered
because of recurrent miscarriages, but others may remain unnoticed. In cases of translocation involving very
small segments of chromosomes, there could be partial trisomies and partial
monosomies in which affected babies could survive till birth, although they
would have congenital malformations.
3.
Inversions
 Inversions
occur when there are two breaks within the same chromosome. The middle segment
is inverted and re-fusion of the broken ends occurs (Fig 6.15).
Inversions
occur when there are two breaks within the same chromosome. The middle segment
is inverted and re-fusion of the broken ends occurs (Fig 6.15).
A peri-centric inversion occurs around the centromere and, as a
result, the relative lengths of the sort and long arms may change.
A para-centric inversion does not involve the centromere. It may
occurs in either the long arm or the short arm., whose lengths are not
affected.
In both types, the
sequence of genes in the inverted segment will be altered. This will cause
problems in pairing during meiosis and may give rise to unbalanced rearrangements in the offspring including duplications and deletions of the
inverted segment.
Some pericentric
inversions do not cause unbalanced rearrangements. An example is pericentric inversion of chromosome 9, which occurs
as an inherited variant.
4. Partial monosomies and partial trisomies
Partial monosomy is the same as a deletion and
means that a segment of chromosome has been lost. There is a single copy of
genes in that segment, instead of a pair.
Partial trisomy occurs if there is an extra copy of a segment of
chromosome in addition to the usual pair.
The genes in that segment are present in triplicate. Partial monosomies and partial trisomies
often occur together. They may be the
result of a balanced translocation or inversion in one of the parents. In some cases they arise de novo, meaning that the unbalanced rearrangement arose in
that particular individual.
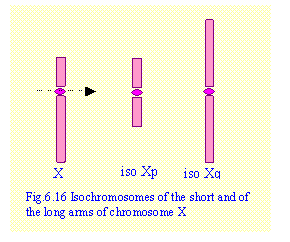 5. Isochromosomes
5. Isochromosomes
Isochromosomes arise if the centromere splits
transversely instead of longitudinally (Fig.6.16). As a result the daughter chromosomes consist of either two long
arms (isochromosome q) or of two short arms (isochromosome p). Isochromosomes are rare. They usually affect the X chromosome. In a
female with isoXp (isochromosome of the
short arm of the X chromosome) there is monosomy of the long arm and trisomy of
the short arm. In isoXq there is
monosmy of the short arm and trisomy of the long arm .
6. Ring chromosome
A ring
chromosome forms when the tips of the long and short arms fuse together to form
a ring  (Fig.
6.17). Usually there are small
deletions of the tips of both arms.
(Fig.
6.17). Usually there are small
deletions of the tips of both arms.
CLINICAL
FEATURES OF CHROMOSOME ANOMALIES
Chromosome
aneuploidies and unbalanced chromosome rearrangements are associated with well-defined
clinical syndromes. A syndrome is a
group of clinical features that are characteristic for a particular
anomaly. Chromosome syndromes have been
described for all chromosomes. Only a
few of the commonest syndromes are outlined here.
Trisomy 21 - Down's Syndrome
Karyotype: 47,XX+21 or 47,XY+21 /
translocation trisomy 21 e.g.
46,XX -14 + t(14;21)
Frequency :
1 in 600 births
Clinical features:
* Mental
retardation
* Hypotonia
* Flat
occiput (brachycephaly)
* Round
flat face
* Slanting palpebral
fissure
* Broad
nose
*
Protruding tongue
* Short
fingers (brachydactyly)
* Increased
frequency of congenital heart disease
Trisomy 18 - Edwards Syndrome
Karyotype: 47,XX+18 or 47,XY+18
Frequency:
1 in 8,000 births; Predominance of
females 4F:1M
Clinical features:
* Growth
retardation
* Small
mouth
*
Micrognathia (small chin)
* Flat,
pointed ears
* Short
neck
* Clenched
hands with overlapping fingers
*
Rocker-bottom feet
* Cleft
lip, congenital heart disease and diaphragmatic hernia are common.
Trisomy 13 - Patau Syndrome
Karyotype
47,XX+13 or 47,XY+13
Frequency
of Trisomy 13: 1 in 10,000 births
Clinical features:
*
Microphthalmia
* Cleft lip
and palate (often bilateral)
*
Polydactyly of hands and feet
*
Microcephaly
* Scalp
defect
* Early death
5p monosomy (deletion) - Cri-du-chat syndrome
Karyotype:
46,XX5p- or 46,XY5p-
Frequency :
1 in 50,000 births
Clinical features:
*
characteristic cat-like cry (due to underdeveloped larynx)
*
microcephaly
* round
"moon-like" face
*
micrognathia
*
hypertelorism
Klinefelter Syndrome - 47,XXY
Frequency:
1 in 1,000 males (occurs only in males)
Clinical features:
*
Testicular atrophy and azoospermia
*
Gynaecomastia
* Usually
tall stature
Turner Syndrome - 45,X
Frequency:
1 in 2,500 females (occurs only in females)
Clinical features:
* short
stature
* lack of
pubertal development
*
amenorrhoea
* ovarian
dysgenesis
* low
posterior hair line
* webbed
neck
PROBLEMS
1. How does the chromatin in interphase nuclei
differ from that in chromosomes?
2. What is the importance of histone proteins in
interphase nuclei and chromosomes? What
would happen if the H1 histones were to be removed by chemical treatment? What would happen if all histone proteins
were to be removed?
3. How does
genetic recombination occur? What is
its importance?
4. In the
following table indicate whether each statement is true (+) or false (-)
regarding mitosis, meiosis I and meiosis II.
|
|
MITOSIS |
MEIOSIS
I |
MEOSIS
II |
|
Is preceded by DNA
replication |
|
|
|
|
Separation of
chromatids occurs |
|
|
|
|
Division of the
centromere |
|
|
|
|
Crossing over between
homologous chromosomes occurs |
|
|
|
|
Results in a haploid
set of chromosomes |
|
|
|
|
Daughter cells are
genetically identical |
|
|
|
|
A spindle is formed |
|
|
|
5. A baby girl is born with clinical features of Down syndrome. Her mother is has a balanced translocation between chromosomes 21 and 15. What would be the karyotype of the child and of her mother?
6. If a person is a carrier of a Robertsonian
translocation between chromosomes 15 and 21, what are the possible chromosome
outcomes in the offspring? Indicate
diagrammatically the chromosomes in the parent, the gametes and the offspring.
7. If a
person is a carrier of a Robertsonian translocation between the two chromosomes
21, what are the possible outcomes in the offspring? Indicate diagrammatically the chromosomes in the parent, the
gametes and the offspring.
8. Explain how Turner syndrome and Klinefelter
syndrome are likely to arise. Can these abnormalities arise from a defect in
meiosis in the father or in the mother?