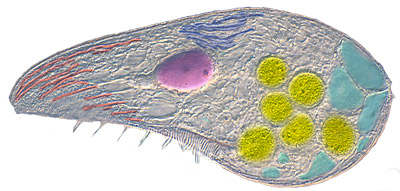
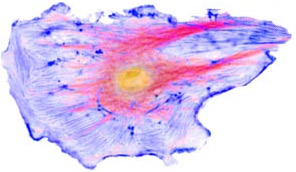
Eukaryotes are complex flexible cells, with various internal compartments performing specialized processes.
100µm
nucleus
endoplasmic reticulum
membranemitochondriavesiclescytoskeleton
Note that free living cells are mostly transparent; cell images are often stained.
The fundamental difference between eukaryotes and bacteria (prokaryotes) is their flexible cellular membrane held together by the cytoskeleton. They have the ability to form interior membrane-enclosed vesicles, some of which are permanent organelles that have specialized functions. Some vesicles form at the endoplasmic reticulum, then sorted by the Golgi body; others form at the cell membrane, to form a two-way traffic between the cell interior and exterior.
Organelles allow synthetic and catabolic processes to occur simultaneously in different compartments. For example, mitochondria are organelles that generate ATP. Concentrating their metabolic reactions in these organelles, eukaryotes can grow bigger, several thousand times the volume of a bacterium (), which in turn makes them less susceptible to osmotic pressure, and so do not normally need a cell wall.
The amount of DNA can be much larger than in bacteria. The nucleus can hold up to bases that code for about proteins (of which are used regularly).
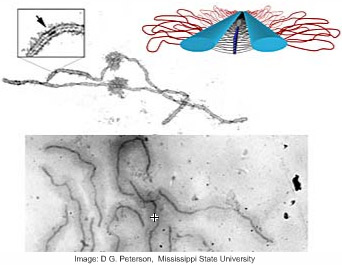
The Cytoskeleton
The cytoskeleton consists of two or three systems of filaments:
MicrofilamentsIntermediate filamentsMicrotubules
A mesh of microfilaments of actin fibers, assembled into helices; several proteins enhance or inhibit assembly, or cap, cut, cross-link or connect them to the membrane. Powered by ATP, they allow the cell to change its shape, hauled relative to each other by the protein myosin, working in tandem. They are found throughout the cell, but especially at the membrane.
There may be other permanent and stronger intermediate filaments (), made up of vimentin or lamin, that fix the cell or vesicle shape.
A spindle of microtubules of tubulin fibers, assembled into tubes growing from a centrosome to the membrane; other proteins can cap, twist or connect them to the membrane. Powered by GTP, they allow vesicles and organelles to move inside the cell, pulled outward or inward by multiple kinesin or dynein protein motors respectively.
Kinesin motors walking along microtubule, dragging vesicles.
Membrane
Cells use NaK-ion pumps, similar to ATP synthase, to pump Na+ out, and keep K+ in; while they pump Cl- out and keep PO42- in; this way there is a weak electric potential across the membrane, while the pH inside is buffered at . In seawater, there is hardly any difference in ion concentration, so osmosis is not a problem, as it is for bacteria. But if the cytoskeleton is damaged, the cell may swell up with water until its membrane ruptures and the cell dies.
Vesicles
Cells use vesicles to transport molecules to organelles and to the outside.
Endocytosis
Vesicles form when a large protein, clathrin, connects with "adapter" proteins on a membrane, and with other clathrins, to assemble a cage with the membrane inside. A second protein then squeezes and cuts the membrane at the neck, and a dynein motor protein carries it inward along a microtubule.
The vesicle traps the exterior solution, as well as any membrane proteins (such as receptors).
Exocytosis
A vesicle fuses with its intended membrane, by attaching to the correct snare protein complexes. Two such matching complexes then merge the two membranes together. The solution inside the vesicle is secreted outside, while the vesicle's membrane proteins become part of the target membrane.
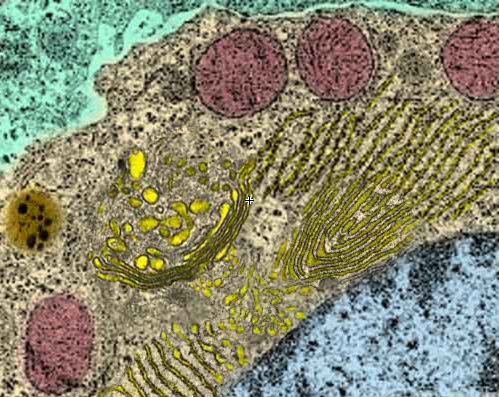
The Endomembrane
Inside the cell, there is a highly folded and interconnected membrane, where membrane lipids and proteins and most carbohydrates are synthesized and sorted. The first three amino acids of a protein are often used as a tag to identify their target organelle.
Smooth Endoplasmic Reticulum: fatty acids, phospholipids, carbohydrates and sterols are synthesized here. It is the source of new membrane.
Rough Endoplasmic Reticulum: when a ribosome forms a polypeptide that is marked to become a membrane protein, an SRP protein attaches to the polypeptide as it is forming, blocking translation, and takes the ribosome-polypeptide to the ER, where it attaches to a receptor; here translation resumes and the peptide becomes embedded in the endomembrane.
Most proteins are transported to their destination inside vesicles, but some molecules are transported inside protein 'vaults', shown here.
The Golgi body sorts out the proteins and packages them into vesicles, each destined to a specific organelle. First, vesicles from the ER, full of lipids, carbohydrates and membrane proteins, fuse together at the proximate end of the Golgi body. Inside, the peptides are modified, e.g., some parts are snipped, some carbohydrates are added to the proteins (now called glycoproteins), adding to their structure and serving to identify their target. Once completed, these proteins sort themselves out by attaching to specific receptors. When enough accumulate, at the other end of the Golgi body, they cause a vesicle to form, marked with a specific address. Motors then drag the vesicle along the microtubules to their target organelle (such as mitochondria, lysosomes or the nucleus) or to the cellular membrane, or back to the ER. Here, the vesicle fuses with its target membrane, although some vesicles may have to await for a specific signal for them to fuse with the cellular membrane and secrete their contents outside the cell.
Cytosol proteins are also marked with the address of their target organelle. They are chaperoned until they reach their destination, where they are imported and the protein folds properly.
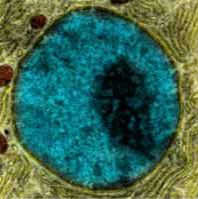
The Nucleus
10µmNucleus with the darker nucleolus, where ribosome RNA is being continually produced, and knots of tightly condensed regions of chromosomes.
The largest 'organelle' is the nucleus which
protects the much longer DNA (), organized into chromosomes, so that they can be repaired and replicated more easily;
separates the transcription of genes (inside) from the translation of mRNA by ribosomes (outside).
In most species of eukaryotic cells, two copies of each chromosome are present (diploidy); thus each gene has two versions (alleles), differing from each other about 1 in 1000 bases. This backup greatly lowers the chance of being affected by a debilitating version of a gene.
Most defective alleles produce a sub/non-functional protein, hence are recessive, overridden by the good allele. For example, if there is a 1 in 10 chance of having a bad recessive allele, then with 2 copies of each gene, the chance of having them both bad reduces to 1 in 100.
But some defective alleles produce a protein that harms the cell, hence are dominant over the other allele, e.g., a transcription factor which permanently switches a gene off, or a polypeptide which sticks badly to another. One copy of a dominant allele is enough to cause disease.
Each gene is found in the same position on the same chromosome, in order to allow the cell to repair a complete double-stranded break in one chromosome using its paired up homologous chromosome (and enable meiosis — see later).
Chromatin
Because chromosomes are so long ( bases), they are
condensed using histone proteins that wrap the DNA in coils, forming a nucleosome for every 147 base pairs.
open ended not looped, with the free ends protected from unzipping by telomeres — a repeating DNA sequence with capping proteins.
The chromosomes' middle section is a repeating sequence of bases, called a centromere, which is attached to the nuclear membrane by proteins, somewhat grouped separately. Most of the DNA in them is highly condensed near the membrane, except those parts that are in current use.
The number and length of chromosomes vary randomly by species. Genes take up only 2—40% of the DNA; the rest is probably safety padding. There are about 15-50 copies of the genes that produce rRNA, tRNA, spliceosomes..., and fewer copies for other essential genes.
Passage of proteins to/from the nucleus is controlled by about 2-3k large protein pores. Ribosomes and mRNA are tagged and actively exported outside where they become functional, while various proteins (histones, repair/replication proteins etc.) are actively imported in (smaller molecules can easily pass through).
mRNA processing
Before transcription can start, a protein complex, called an enhanceosome, forms around the transcription factors; its job is to disassemble the nucleosomes, so that the RNA polymerase can attach to start transcription.
mRNA is processed further inside the nucleus before being exported:
Their ends are protected by proteins and a repeating adenosine sequence is added at one end,
An RNA spliceosome forms at specific points of the mRNA, and cuts out (splices) 0-30 stretches of DNA, called introns; the remaining exons are rejoined, ready for export. The cell can decide which combination of exons to include in the final mRNA, making it possible for one gene to code for multiple variants of proteins, to be used in the different regions (membrane, cytoplasm, or organelles) or cell stages.
Eukaryotes have a more complex regulatory network than bacteria because of the larger number of genes:
there are more non-coding RNA genes than protein genes; these are probably involved in regulation;
there are many more activator/repressor proteins to control enzymes, mRNA;
combinations of transcription factors attach to "enhancer"/"silencer" DNA regions to uncoil the DNA, and switch on/off genes; (10% of genome consists of transcription factors);
methylation of genes is used to switch them off permanently;
mRNA can be quickly switched off when special complementary micro-RNA attach to it; 'dicer' proteins then find and cut up the double stranded RNA, while 'RISC' proteins use the resulting bits of RNA to destroy any other matching mRNA (e.g. viral). Thus a gene can be rapidly silenced (or slowed) well before it is switched off by inhibitor proteins.
Chromosome Breaks
Repair enzymes often manage to reconnect the two ends of a broken chromosome, using the homologous chromosome:
A complete chromosome break occurs...The free ends are capped and a short portion of a strand removed.The end is matched with its homologous chromosome.The strand is extended, copying from the other chromosome.The other free end matches and the final gap filled in.The break is repaired flawlessly.
... but sometimes, things go wrong:
Repair enzymes often manage to reconnect the two ends of a broken chromosome. But if the other end is not found, new telomeres are produced, creating an extra chromosome.
More rarely there are multiple breaks (especially in 'fragile' regions), and the wrong ends may be joined by the repair mechanism; if the strands are on different chromosomes, the result is a translocation or fusion of chromosomes.
If crossing DNA strands on the same chromosome are switched; the result is an inversion or a missing fragment.
If the breaks do not involve genes, there is little effect, but there are serious effects if a gene is broken, or it loses its control region, or a dormant gene is transferred to an active region. Even worse, deletions of a chromosome (>1Mb) expose the bad alleles of the other chromosome, hence the cell usually dies. If chromosome number/form changes, reproduction cannot occur since the gametes that are produced are unable to perform meiosis as the chromosomes cannot pair up.
Mutations
The DNA of two individuals in a species differ in about bases. With time, rare DNA replication errors accumulate, some of which spread by inheritance — about 1 base every generations.
Of all the mutations,
80% are inserts ('satellite' variations),
20% single base changes (called SNPs)
of these, only 1/1000 are in an exon of a gene;
of these only a 1/3 produce non-synonymous amino-acids,
of these, 1/100 are sufficiently different to cause disease.
After 250k generations, as mutations accumulate, there is enough difference from the original to make a new species; after a million generations, the difference is bases from the original, equivalent to a new genus.
Over evolutionary time, chromosome reconnections and duplications of gene families also lead to the genome being shuffled, about one event every 150k generations; hence species diverge with different chromosome length and number.
This simulation traces the frequency of two alleles A and B of a single gene:
A:B
Generations
AA
AB
BB
0.991.01
Fitness advantage
Genetic Drift: If AA = AB = BB in fitness, then the alleles A,B vary randomly.
Rare Disease: If AA = AB > BB in fitness, then the B allele dies out slowly, kept in the genetic pool by the healthy combination AB.
Recessive Advantage: If BB > AB,AA, it is hard for the (novel) recessive allele B to increase, but once it gets a foothold, it replaces the A allele.
Heterozygous Advantage: If AB > AA,BB, then both alleles are kept indefinitely in the genetic pool.
Heterozygous Disadvantage: If AB < AA,BB, then one alleles eventually wins out.
Organelles
Phagocytosis and the Lysosome
Various heterotrophic eukaryotes engulf other lives/organic material by maneuvering their cytoskeleton to surround them, guided by receptors. The resulting 'vacuole' is merged with one or several lysosomes.
Lysosomes are large vesicles formed by the fusion of smaller ones. They contain membrane enzymes that attack peptides, nucleic acids, lipids and carbohydrates. Upon fusion with a vacuole, proton pumps make it acidic inside, activating the enzymes, e.g., lysozyme attacks bacterial cell wall. Most organic material is quickly degraded and absorbed into the cell. The vacuole then merges with the cell membrane, expelling whatever remains inside.
The ability to engulf bacteria has led to several symbiotic/slave relations.
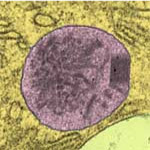
Peroxisome
2µm
Peroxisomes are large vesicles with membrane enzymes that perform oxidative and anti-oxidant reactions.
Some oxidize fats to shorter soluble pieces that are exported out of the vesicle, reducing oxygen O2 to hydrogen peroxide H2O2 in the process. Catalase then gets rid of the dangerously reactive peroxide by converting it back to O2. They also mark various 'unwanted' molecules (poisons) with glutathione, to be exported out of the cell.
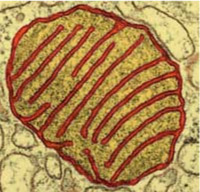
Mitochondria
1-10µm mitochondrion with its internal voluted membrane
Mitochondria are bacteria-sized organelles that carry out most of the energy-producing catabolic reactions; they provide ATP for the cell. Each cell has several of them, depending on how active it is.
Like their bacterial ancestors, they import pyruvate, G3P, fatty acids and ketones from the cell, and break them down to acetate, which is used in the citrate cycle to produce NADH. Membrane protein complexes oxidise it with oxygen, in the process producing an electric potential across the inner membrane. ATP synthase then uses this energy to regenerate ATP, that is then exported back to the cell.
Malate and citrate freely move in and out; the latter is used by the cell to inhibit glycolysis.
Mitochondria are also involved in the production of amino-acids: they aminate (and deaminate) ketoglutarate to glutamate, oxaloacetate to aspartate, ammonia to glycine and carbamoyl, ornithine to citrulline.
They have their own small amount of DNA (0.1M bases, 37 genes) that code for the essentials: ribosomes, tRNA, ATP synthase, cytochromes, ... They divide by fission when signalled to do so.
Entry and exit to the mitochondria is controlled by proteins in both the inner and outer membranes. Proteins destined for them are kept unfolded by small chaperone proteins until they reach a receptor in the outer membrane. Here the protein tag is cut off, and the protein imported and folded.
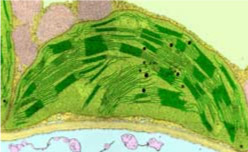
Chloroplasts
A chloroplast, 3-10µm, with its interconnected internal stacks of disk-shaped grana.Photosynthetic eukaryotes have chloroplasts, which absorb light to produce ATP and NADH. Chloroplasts are full of rubisco — the most plentiful enzyme of all — to absorb CO2.
Protein photosystems are found at the surface of the grana. Their chlorophylls absorb red and blue light and use its energy to oxidize water into oxygen, pumping protons out into the grana. In the process, they also reduce NADP to NADPH. A third membrane protein, ATP synthase, then uses the protons to produce ATP from ADP.
The high levels of NADPH and ATP are used to produce and export 3-carbon molecules (G3P, PGA...) and to synthesize starch, a polymer of glucose.
As with mitochondria, proteins destined for chloroplasts are tagged, kept unfolded by chaperones, and then imported inside them by membrane ports.
Chloroplasts are remnants of cyanobacteria, and still retain less than 100 genes.
Contractile Vacuole
A spherical contractile vacuole; some species have more complex and permanent vacuoles with tubes.
Eukaryotes that live in fresh water absorb water slowly through osmosis. They eliminate this extra water by forming an internal vacuole; pure water enters it passively; when it grows, in less than a minute, it is merged with the cell membrane, expelling the water.
Land plants have a modified permanent 'sap' vacuole; it stores the water and keeps the cell pressure up.
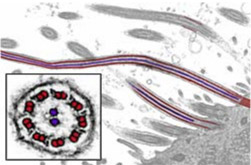
Flagella
0.2-0.5µm
Eukaryotes often have one or several flagella. At their base, a centriole grows nine pairs of microtubules anchored around a central pair. Dynein motor proteins can pull the outer tubules relative to each other, causing the flagellum tip to bend in rapid wave-like fashion, pulling the whole cell forward at several cell-lengths per second.
Some eukaryotes have several short flagella, called cilia, that beat in synchrony, making them even faster.
There may be a light-sensitive pigment controlling the flagella.
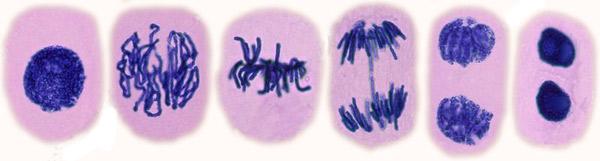
Cell Division: Mitosis
To replicate and separate a billions-long DNA genome is a delicate and lengthy affair.
G1, S, G2ProphaseMetaphaseAnaphaseTelophase
G1 As the cell grows, it prepares itself for replication. It
produces more ribosomes and the required proteins, and more organelles;
performs DNA repair and checks that there are no major faults such as double strand breaks.
This may take up to 9 hours and is strictly regulated. Once a cell commits to start dividing, there is no turning back.
S-phase When ready, the histones and gene promoters/inhibitors are removed, and the DNA replicates slowly at (with short reverse-strand fragments of 100-150 bases); there are multiple origins (about every 50k – 300k bases) and several (20 – 80) initiator proteins, taking 7-15 hours to finish the job. After DNA replication, a special protein scans the DNA looking for methylated bases on one strand only and adding a methyl group to the other strand (so switched-off genes remain so after mitosis). When a DNA replisome reaches the free end of a chromosome, the last adenosine base of the 'reverse strand' cannot be replicated, and is omitted; the telomere therefore shortens with each replication, unless it is lengthened periodically by a telomerase protein.
G2 The cell continues to repair its DNA, using both chromosomes for accurate copying.
Prophase In 5 hours, the chromatin is condensed into paired chromatids, joined at their centromere. Transcription stops completely. The lamin cytoskeleton disassembles, fragmenting the nucleus and endomembrane into small vesicles. Meanwhile, the cell's centrosome replicates and are pulled to opposite ends.
Metaphase The centrosomes form the mitotic spindle: microtubules grow from each centrosome until they meet either the cell membrane, the opposite tubules or a centromere. Two kinetochore protein complexes attach to the centromeres of the joined chromatids and pull at the tubules. This brings a chromosome pair to the center of the spindle in about 20 mins.
Anaphase When all the chromosome pairs are aligned in the middle, a trigger causes the splitting of the centromere proteins joining each pair. This frees the kinetochore motors to pull each chromosome towards the centrosomes, which they reach after about 20 mins of pulling. The centrosomes are also pulled further towards the cell membrane.
Telophase When separation is over, the spindle disassembles, and the endomembrane reforms around each chromosome, joining up until two separate nuclei form; the chromosomes uncoil and resume transcription. The cytoskeleton reforms, some actin forming a ring in the middle whose filaments move past each other causing it to contract. This pulls the cell membrane with it, splitting the cell in two.
The most common (but rare) error in mitosis is that a paired up chromosome fails to separate. As the cell divides around it, one daughter is left crippled without the chromosome and soon dies, while the other has two copies and is only slightly at a disadvantage to normal cells. Very rarely, the separation process is arrested, and the cell ends up with 4 copies of each chromosome; it dies.
Cell Fusion
Meiosis
The flexibility of the cell membrane allows for the possibility of fusing cells and mixing chromosomes among different cells via gametes. Having hybrid daughter cells with novel combinations of genes, increases the chances of survival of one of them in a harsh environment, or against parasites. This means that genes now compete in a gene pool of alleles, speeding up adaptation.
Meiosis takes much longer than mitosis. The chromosomes first replicate. Then each of the two corresponding ('homologous') chromosome pairs align parallel to each other, held together by 'zip'-proteins.
At random hotspots on the chromosome pairs, a crossover ('chiasmata') of DNA occurs: about one hotspot every bases; one crossover every hotspots, i.e., 1-3 every chromosome. Effectively, the 4 chromatids are randomly reshuffled between themselves. This is only possible because homologous chromosomes have the same genes in the same positions.
The zip-proteins then dissolve, the chromosomes condense, and mitosis separates them into two daughter cells, with homologous chromosomes still attached at the chiasmata.
A second mitosis separates the homologous chromosomes, forming 4 hybrid gametes, each with a set of chromosomes, genetically different from the parent cell.
Rarely, the paired up chromosomes fail to separate: one gamete ends up with a missing chromosome, while the other has two. More rarely, (1 in a million) chromosomes pair up at misaligned points during cross-over; then one daughter chromatid loses a fragment (deletion) to the other (duplication).
Fertilization
When two gametes meet, one of them dissolves part of the other's membrane, becoming one cell. Eventually, the two nuclei fuse together, forming a diploid zygote. To avoid self-fertilization (which certainly doubles the faulty alleles), eukaryotic cells have a type, + or -, of membrane receptor; gametes with the same receptor (such as from the same parent) do not fuse.
Rarely, three gametes fuse (about 1 in 50 chance), or a gamete fuses with a diploid cell (1 in 100). The result is a cell with 3 copies of each chromosome; it usually dies.
Life-Cycle
Whether a eukaryotic cell spends most of its life with a single (haploid) or double (diploid) the number of chromosomes depends on the species.
Originally, eukaryotic cells only fertilized just before forming a cyst, in stressful times when variation of cells is at a premium.



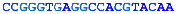





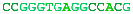
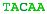
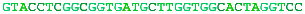
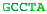

 A complete chromosome break occurs...The free ends are capped and a short portion of a strand removed.The end is matched with its homologous chromosome.The strand is extended, copying from the other chromosome.The other free end matches and the final gap filled in.The break is repaired flawlessly.
A complete chromosome break occurs...The free ends are capped and a short portion of a strand removed.The end is matched with its homologous chromosome.The strand is extended, copying from the other chromosome.The other free end matches and the final gap filled in.The break is repaired flawlessly.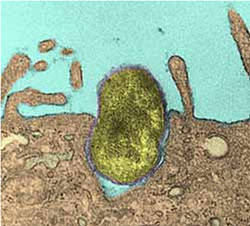

 G1, S, G2ProphaseMetaphaseAnaphaseTelophase
G1, S, G2ProphaseMetaphaseAnaphaseTelophase